

На Всероссийской научной конференции «Творчество в современном мире: человек, общество, технологии», проведенной Институтом психологии Российской академии наук, с докладом «Эволюционный процесс на индивидуальном и социокультурном уровне» выступил Юрий Александров.
Юрий Иосифович Александров – доктор психологических наук, профессор, член-корреспондент РАО, заведующий лабораторией психофизиологии им. В.Б. Швыркова ИП РАН, заведующий кафедрой психофизиологии ГАУГН, заведующий лабораторией Нейрокогнитивных исследований МГППУ, профессор ВШЭ.
«Центральным пунктом в исследовании системно-эволюционной парадигмы является память и научение. Почему? Потому что в результате научения, которое ведет к формированию памяти, возникают довольно сложные структуры опыта. И актуализация этих структур, их активация может быть описана психологически, физиологически, молекулярно-биологически и с помощью многих других описаний. Эти описания являются вторичными по отношению к структурам опыта.
Эти структуры и их нейрокогнитивное, психологическое описание находятся в хорошем соответствии с теоретическими представлениями Якова Александровича Пономарева: ЭУС = этапы – уровни – ступени. Этот закон – принцип трансформации этапов развития явления в структурные уровни его организации. Закон ЭУС, как считал Яков Александрович (и я с ним солидарен), может быть применен к анализу живых систем любой стадии развития.
Каким образом он воплощается в наших представлениях? Основой наших разработок, современной альтернативой инструктивным теориям научения являются селективные теории научения. Два наиболее важных, хотя и не единственных, творца этих теорий – Джеральд Эдельман с его нейродарвинизмом и Вячеслав Борисович Швырков с системно-селекционной теорией научения. Они разработали эти представления независимо друг от друга. Центральными пунктами селектвных теорий являются селекция и последующая специализация нейронов в отношении вновь формируемых действий. Селекция, это очень важно, предполагает разнообразие нейронов вместо рассмотрения их в качестве гомогенных узлов нейросетей.
Каким образом формируется системная структура субъективного опыта? Составляющие ее элементы – системы формируются на последовательных этапах развития индивида в процессе системогенеза –научения. В отношении этих систем специализируются определенные группы нейронов, причем под каждую систему в процесс специализации вовлекаются новые группы нейронов, отличные от тех, что специализировались ранее в отношении других систем. Это означает, что нейроны – системоспецифичны. Таким образом получается, что когда новые нейроны формируют свою специализацию, они как бы прибавляются к уже специализированным и, таким образом, оказывается, что вновь образованные системы «наслаиваются» на предыдущие, ранее образованные системы – слои опыта (см. рис.1). Эволюция необратима, принципиальной является необратимость индивидуального развития. Особенно четко это было сформулировано И.Р. Пригожиным, он считал, что необратимость - механизм, который создает порядок, организацию из хаоса.
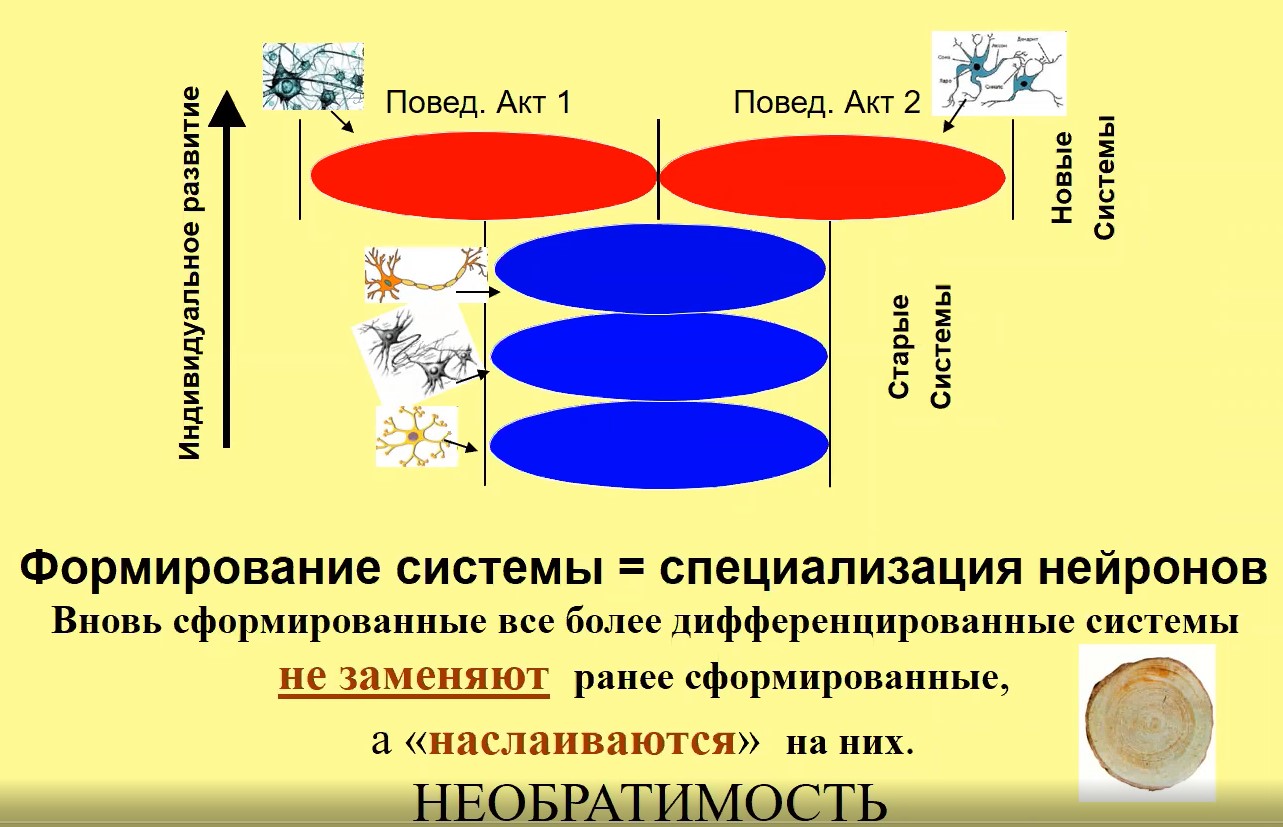
Рис.1
Каким образом формируется память? В начале формирования памяти находится рассогласование. Это рассогласование может быть рассмотрено как на индивидуальном, так и на клеточном уровне. На индивидуальном оно означает противоречие между необходимостью достичь положительный результат и отсутствием соответствующего опыта достижения. На клеточном уровне рассогласование – это несоответствие между метаболическими «потребностями» клетки и наличным метаболическим притоком; результатом рассогласования является активация «ранних» генов (регуляторных), а потом «поздних» генов (морфогенетических). Активация поздних генов ведет к изменению морфологии клетки в процессе научения.
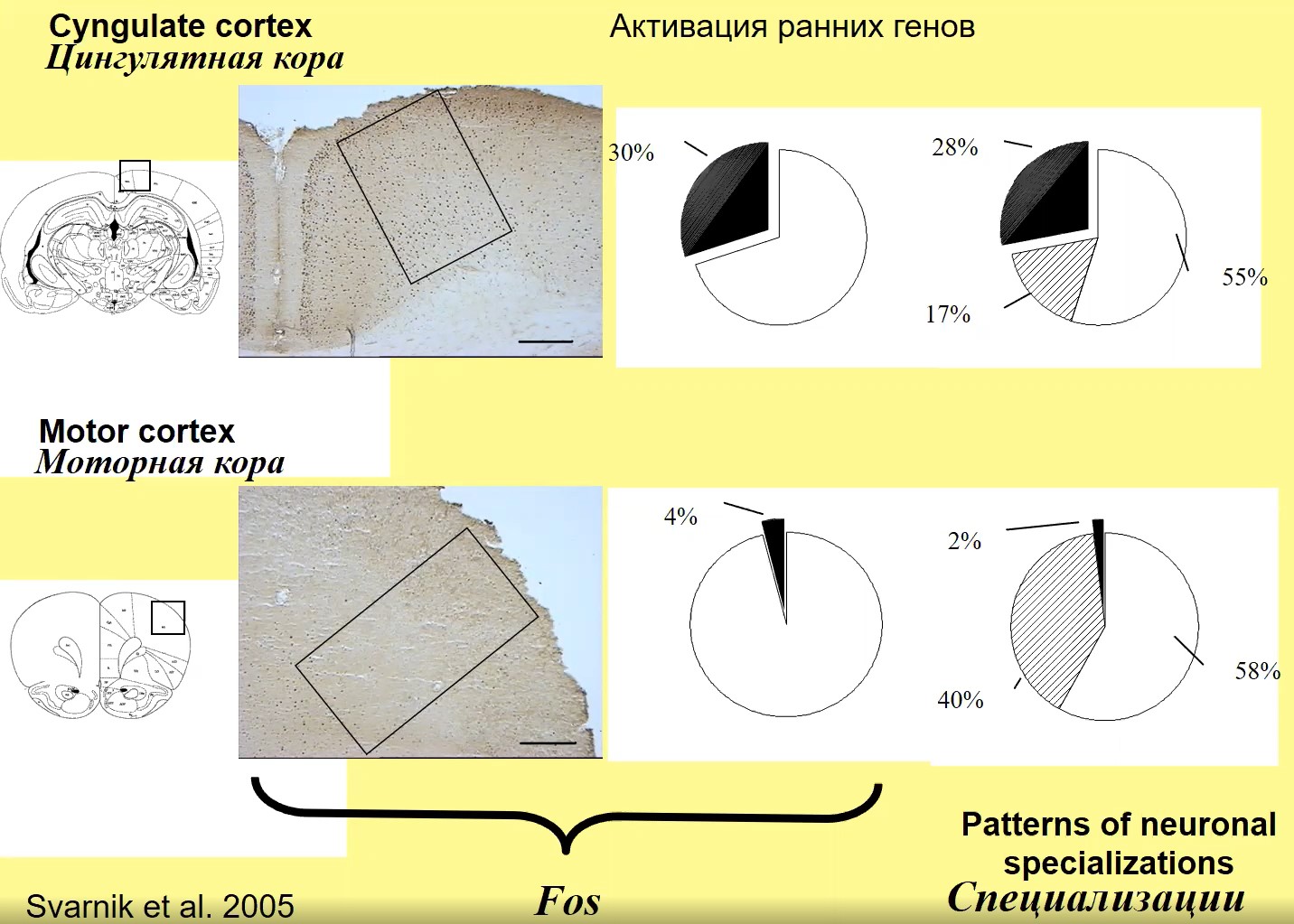
Рис.2
На рис. 2 можно видеть, как выглядят при иммуногистохимическом анализе экспрессия ранних генов. Видно, что этой экспрессии много в той структуре (цингулярная кора), где потом много нейронов специализируется в процессе научения. И пропорционально меньше активирующихся генетически нейронов там, где потом меньше специализируется клеток (моторная кора).
Принципиальная вещь – сначала из массы нейронов, которые есть в мозгу, селектируется на первой стадии определенная группа, в которой возникает экспрессия, и затем из тех, кто генетически активировался во время второй волны селекции только меньшая часть – около 1%. Это те клетки, которые потом будут специализированы. Таким образом специализируется лишь 1% из нейронов, которые генетически активировались для последующей стадии специализации.
И в концепции Швыркова, и в концепции Эдельмана, если рассматривать жизнь индивида, можно выделить первую стадию селекции – это формирование первичного ассортимента разных групп клеток (Эдельман), в наших представлениях это называется «пре-специализации нейронов». В процессе этой селекции наблюдается массовая гибель клеток, т.е. те клетки, которые не сформировали этот первичный ассортимент потенциально специи ализированных клеток, гибнут. На второй стадии из этого первичного ассортимента отбирается вторичный ассортимент, т.е. клетки, специализируемые относительно конкретных поведенческих актов, формируемых в реальной взрослой жизни индивида. При этом, Эдельман отмечает, что селекция №2 у взрослых не сопровождается гибелью клеток. Как показывают наши исследования, проведенные совместно с лабораторией В.В. Шерстнева, на самом деле при селекции №2 также наблюдается гибель клеток, т.е. у взрослого человека также клетки гибнут, у них развивается альтруистичный суицид – апоптоз, который сопровождает формирование специализации у выживших клеток.
При этом у них меняется морфология.
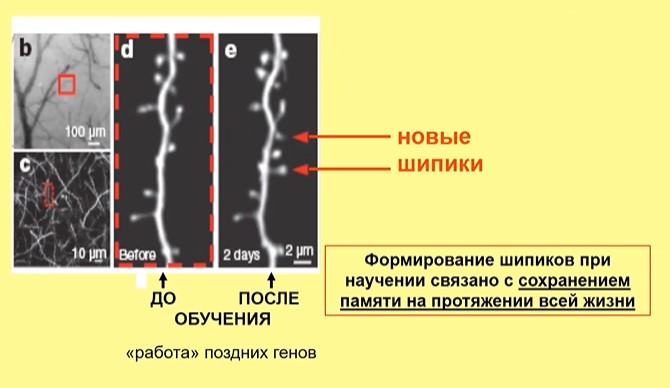
Рис.3
На рис.3 результаты прижизненной регистрация морфологии дендритов нейронов у свободно-подвижного животного. Видно, что появились в результате обучения новые шипики. Формирование этой морфологии связывается авторами с сохранением памяти на протяжении всей жизни.
Мы показали в массе экспериментов, что специализации нейронов формируются не относительно функций (эмоций, сознания, памяти, внимания и т.д.), а относительно конкретных результатов в соотношении индивида и среды. Положение Якова Александровича Пономарева: «Память выступает как условие хранения субъектом результатов», с точки зрения этого эмпирического материала, является абсолютно верным. Связь активности специализированных нейронов с достижением результатов феноменологически выражается в том, что внешне одинаковое поведение, например, нажатие кнопки человеком, будет обеспечиваться разным набором активирующихся специализированных нейронов, в зависимости от того, какая цель данным нажатием достигается. Здесь мы сталкиваемся с межсистемным разнообразием клеток: в разных поведениях активируется специфический набор клеток.
Элементы, которые входят в систему, направленную на получение результата, не просто взаимодействуют, т.е. активируют друг друга, они взаимоСОдействуют. Их активность скоррелирована для достижения результата этой системы – соотношения целого организма со средой. Активность клеток, в том числе нейронов, направлена на обеспечение этого взаимосодействия, результат которого на клеточном уровне – это достижение метаболического результата каждой клеткой.
Различие между клетками обусловлено потребностями метаболической кооперации. Клетки не только разные, но комплементарно (взаимодополнительно) разные. Достаточно обратиться к тому, что известно, например, об отношении нейронов и глиальных клеток, там будет сразу видно, как метаболические циклы одной клетки зависят от метаболических циклов другой клетки и одни не могут существовать без других.
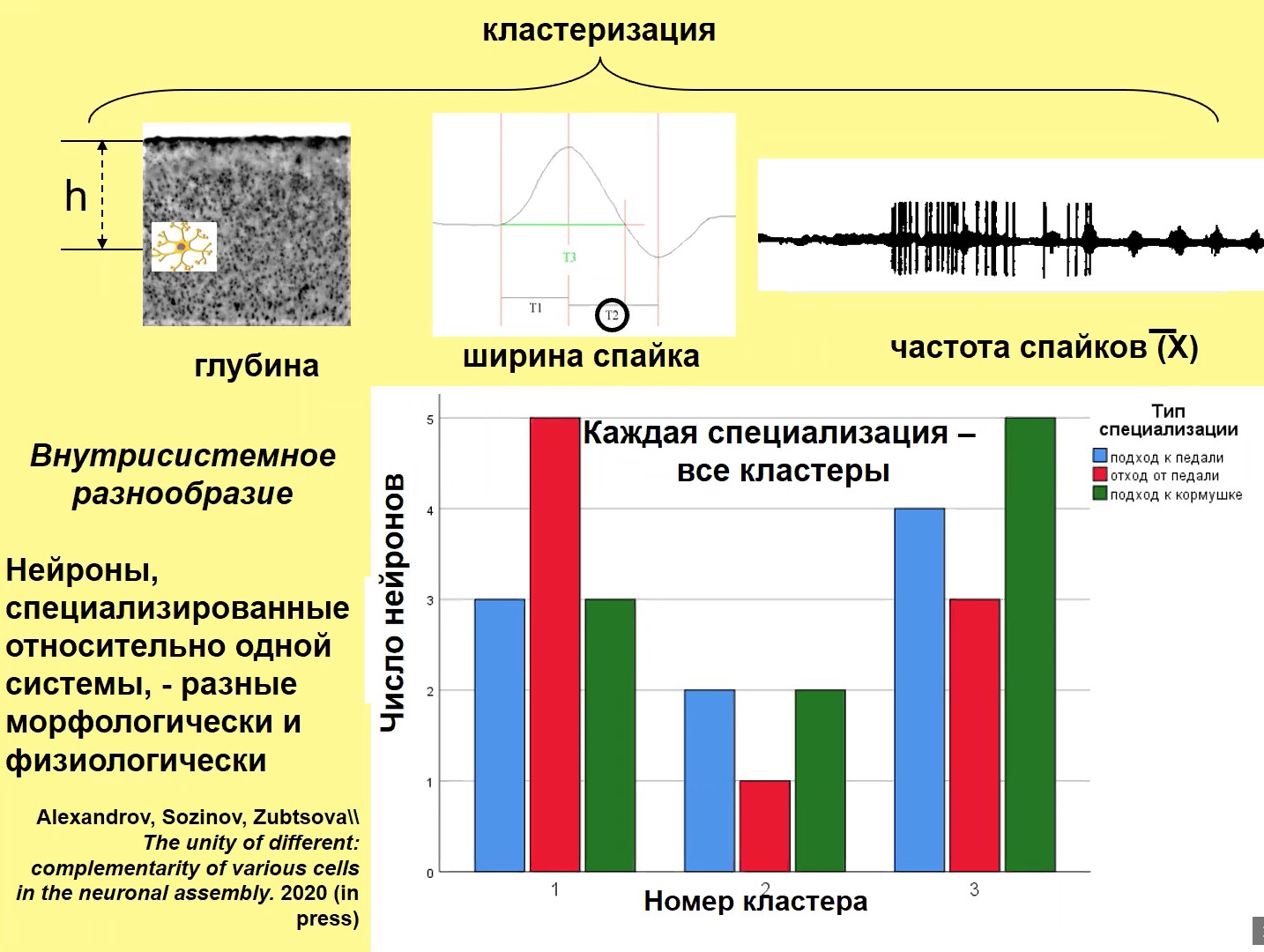
Рис. 4
На рис. 4 разнообразие нейронов представлено в несколько ином смысле: речь идет не о межсистемном, а о внутрисистемном разнообразии. Мы исследовали клетки одной специализации (например, клетки, которые обеспечивают достижение конкретного результата – подход к педали, инструментальное нажатие которой обеспечивает получение пищи). Мы изучали, на какой глубине они находятся, к какому слою коры соответственно они принадлежат, какова ширина спайков у этих нейронов (ширина спайков – физиологический параметр, отражающий также и морфологические характеристики нейронов), какова частота спайков. Мы кластеризовали эти данные. Оказалось, что каждой специализации принадлежат нейроны, принадлежащие к разным кластерам, т.е. различны морфологически, физиологически. Иначе говоря, состав каждой системы – гетерогенный.
Это «комплементарное разнообразие» – эволюционная находка, воспроизводимая на всем протяжении эволюции и на всех уровнях организации.
Например, при изучении микробных колоний, микробных сообществ гетерогенность связывается с адаптацией колонии. Эти микроорганизмы достигают совместных результатов как оборонительных, так и пищедобывательных или дыхательных. Их (клеток) гетерогенность связана как раз с необходимостью реализации разных степеней свободы для обеспечения этих результатов.
Известны также данные о межвидовой комплементарности. Межвидовая комплементарность обеспечивает лучшую адаптацию и лучшее выживание каждого из комплементирующих видов в природе.
Мы можем видеть рост разнообразия (сложности) как в филогенезе, так и на уровне отдельного организма, в онтогенезе.
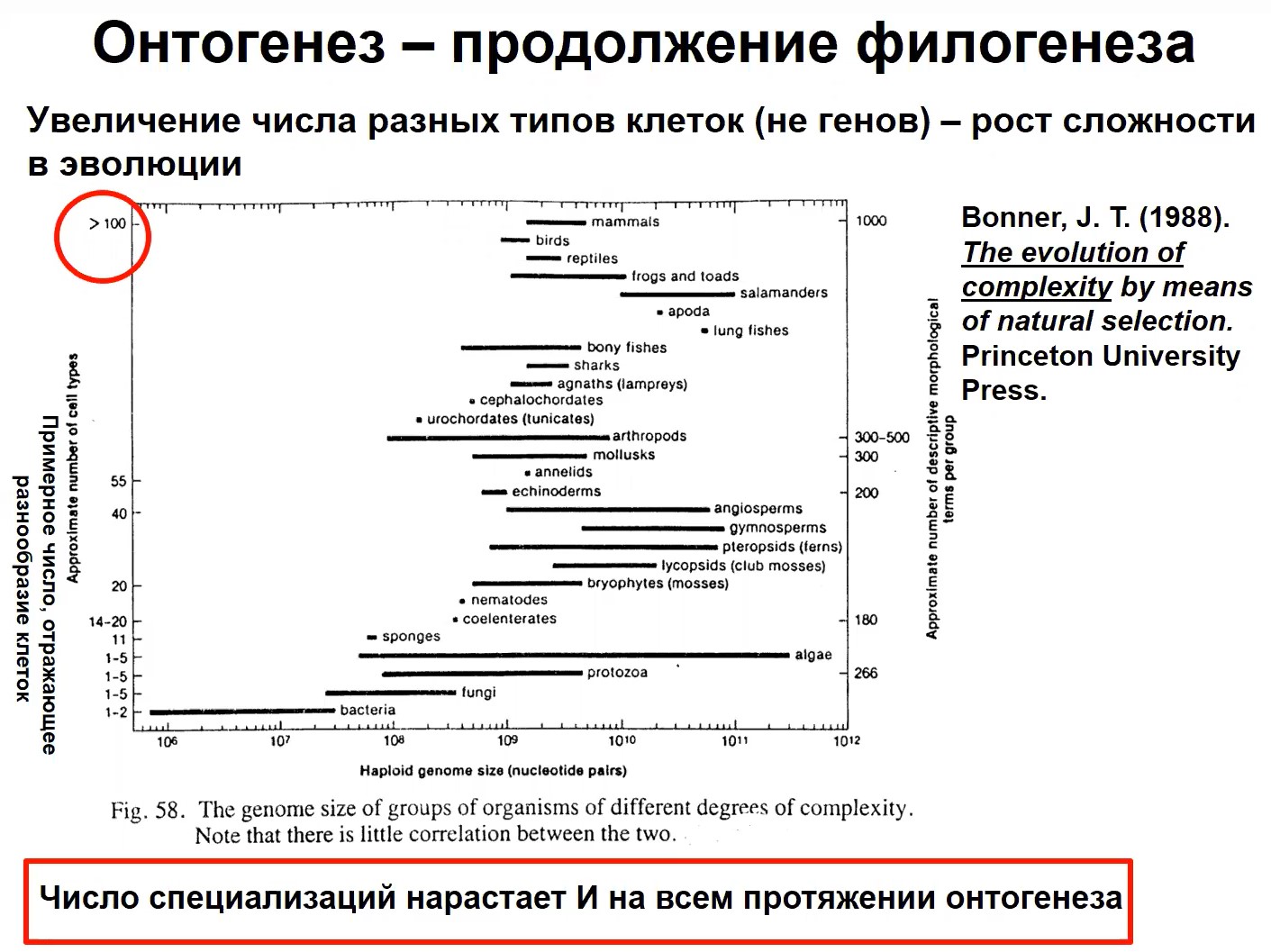
Рис.5
В таблице, приведенной на рис. 5 и взятой из работ Бонера, показана эволюция сложности посредством селекции. Эта эволюция сложности от бактерий до млекопитающих не коррелирует с величиной генома (по абсциссе). Она коррелирует с числом различающихся типов клеток у организма (по ординате): чем больше клеток, тем больше сложность организации, тем более сложные организмы. Заметим, что число специализаций нарастает и во время онтогенеза. Я уже говорил о том, что клетки специализируются, и морфологически, и активационно их различие возрастает на протяжении онтогенеза и число специализированных клеток растет весь онтогенез. Неудивительно поэтому, что как некоторые авторы демонстрируют, сложность электроэнцефалограммы растет на протяжении онтогенеза от ребенка к пожилому человеку. Авторы отмечают, что сложность электроэнцефалограммы отражает все увеличивающийся набор ансамблей нейронов с различающейся специализацией, которые образуются в процессе развития.
Системы – не только общемозговые, но и общеорганизменные. Т.е. для того, чтобы достигать результата взаимодействия со средой, нужны не только нейроны, но и клетки почек, легких, сердца, крови, эпителия, желез и т.д. Все вместе они соорганизуют свою активность и способствуют достижению результата целостным организмом, а каждая из клеток получает те метаболиты, которые ей нужны. Таким образом, комплементарное разнообразие не ограничено только нейронами, а касается вообще клеток организма.
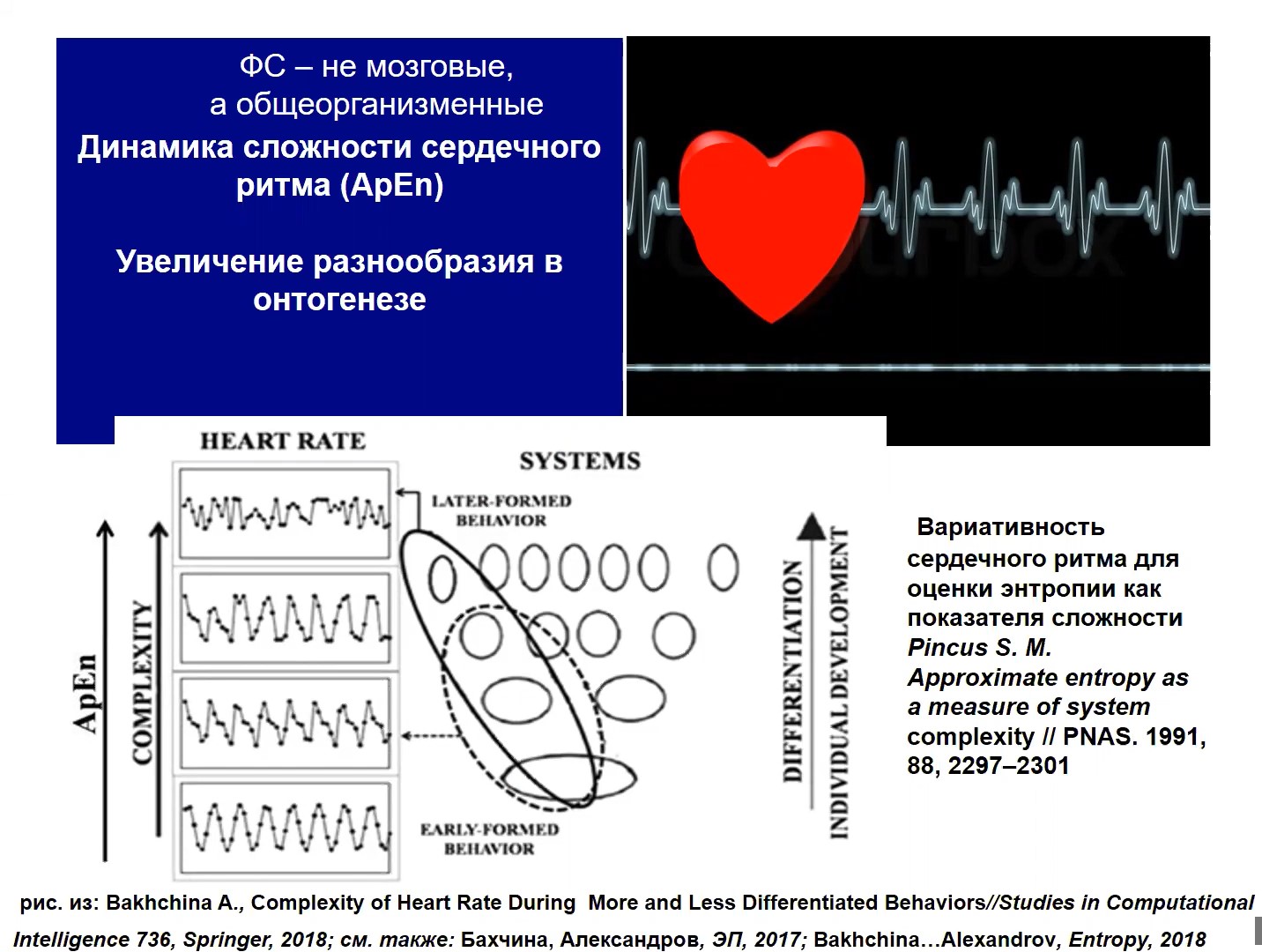
Рис.6
Рост комплементарного разнообразия (сложности) прекрасно демонстрируется при изучении сердечной деятельности (рис. 6). Когда мы изучаем вариабельность сердечного ритма с помощью анализа выборочной энтропии, то мы видим, что сложность сердечной деятельности растет вместе с тем, как все более и более сложные, дифференцированные системы активируются из состава субъективного опыта. Т.е. чем более сложное поведение мы реализуем, чем более сложный опыт актуализируется, тем более высокую энтропию демонстрирует сердце.
Яков Александрович отмечал: «В широком смысле слова память сливается с психикой. Генезис развития психики есть вместе с тем генезис развития памяти».
Это утверждение абсолютно соответствует системно-эволюционному решению психофизиологической проблемы. Корни этого решения находятся в двухаспектной теории, в идеях Гегеля, в идеях Спинозы, в идеях Бехтерева. Системные процессы могут быть описаны и с психологической, и с физиологической стороны – они являются аспектами описания единой системной реальности.
Пример психологического описания дан в единой концепции сознания и эмоций, которую мы довольно давно сформулировали. Здесь я опять обращаюсь к Якову Александровичу, к его утверждению о том, что факт сближения с благоприятствующим, удаления от разрушающего – этот факт специфического ориентирования живого существа по отношению к окружающему мы и рассматриваем, – писал он, – как отправной, кардинальный факт. Это был и для нас один из важных пунктов построения единой концепции сознания и эмоций.
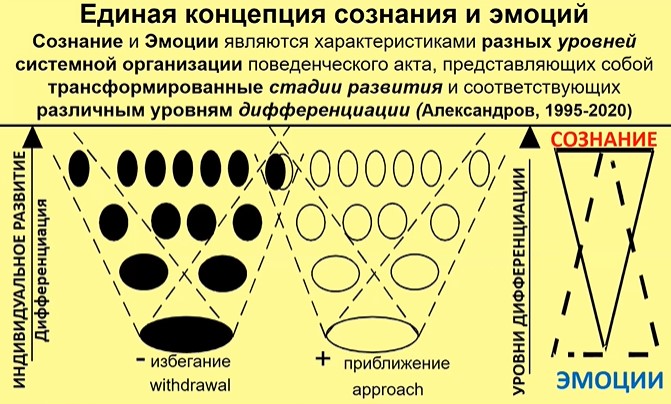
Рис.7
На рис. 7 изображены супердомены опыта, связанные с избеганием (отрицательные эмоции) и с приближением (положительные эмоции). В этих доменах все разное – дианмика формирования памяти, наборы вовлекающихся клеток. Имеются и некоторые перекрытия, как отражено на рисунке. Сознание и эмоции здесь характеризуют разные уровни дифференциации систем. Эти системы формируются на последовательных этапах развития, оно становится все более сложным и более дифференцированным. И, что для нас здесь принципиально важно, эмоции преимущественно характеризуют актуализацию «старых» систем и соотносимы со сравнительно низко дифференцированными уровнями организации поведения.
«Формирование нового опыта соотношения со средой начинается с интуитивного типа взаимодействия и заканчивается рациональным», – для Я.А.Пономарева это было совершенно принципиальное утверждение. Как оно работает, например, в области исследования стресса?
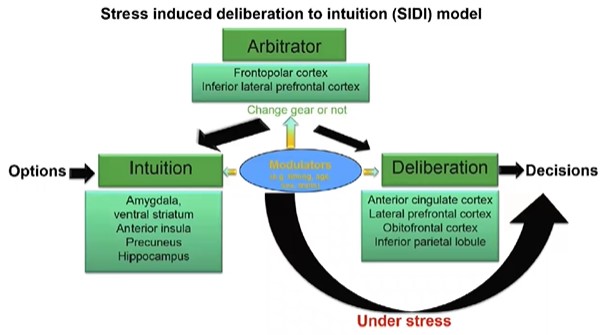
Рис.8
На рис. 8 представлена суммация представлений о стрессе: при стрессе имеются механизмы «перепрыгивания», т.е. переход от интуиции к решению, минуя обдумывание. Каковы механизмы этого «перепрыгивания»? Мы специально анализировали множество состояний (эмоции, стресс, научение, прием алкоголя, болезни) и обнаружили, что у всех этих феноменов есть общий механизм – это механизм регрессии, возврат в прошлое.
Каковы механизмы этого «возврата в прошлое», в «детство»? Это во всех случаях блокирование наиболее сложных, наиболее дифференцированных систем, переход к более простому, «эмоциональному поведению». Отметим, что это не приходит в рассогласование с утверждением о необратимости: это, во-первых, временный процесс блокирования, во-вторых, «полного детства» не наступает.
Экспериментальной демонстрацией этой регрессии (см. рис. 9) является возвращение к «детским» формам поведения при решении моральных дилемм: взрослые без стресса не поддерживают своего, если он не прав, но в состоянии стресса они начинают поддерживать своего даже в случае, если он не прав. Эта поддержка становится такой же, как у детей 3-7 лет.
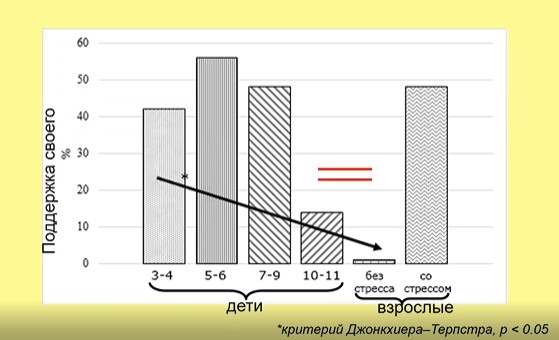
Рис. 9
Когда мы изучаем активность сердца, энтропию, то мы обращаем внимание, что в ситуациях регрессии, де-дифференциации уменьшается, естественно, энтропия сердечной деятельности. Например, при стрессе, связанном с публичным выступлением, энтропия сердца падает (рис.10).
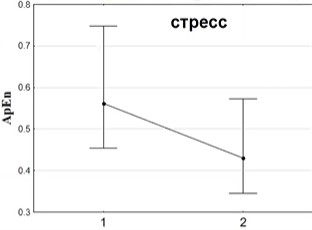
Рис.10
У нас имеются начальные данные, полученные А.В. Бахчиной и соавторами в пользу того, что падение системной сложности, падение дифференциации имеется и в начале обучения, когда надо решить сложную проблему, а решение еще отсутствует (рис. 11). Контринтуитивная вещь: вместо того, чтобы со страшной силой усложняться, индивид как бы упрощается.
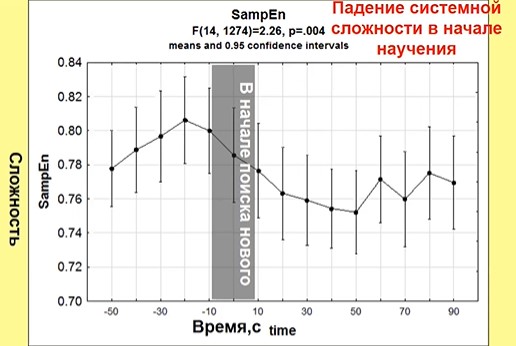
Рис.11
Построение математической модели А. К. Крыловым показало, что модельные агенты, у которых моделируют де-дифференциацию (регрессию), значительно быстрее достигают высокой результативности поведения и находят новые пути, чем те, у которых регрессии не происходит (рис. 12).
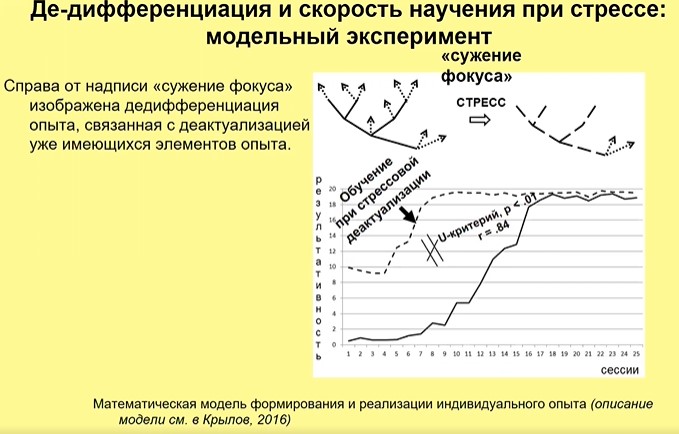
Рис. 12
Мы делаем вывод, что регрессия может достоверно ускорять научение в новой для индивида проблемной области за счет того, что «отключается» опыт («не подходящий» для решения новой задачи), и решение «фокусируется», сразу обращаясь к поиску новых для индивида способов.

Рис. 13
Этот рисунок (13) изображает идею, метафору К.Г. Юнга: чтобы подняться на новую вершину знания, надо сперва спуститься в долину. Его позиция, с которой согласуется наша и наши данные, состоит в том, что «регрессия есть основное условие творческого акта».
Должен сказать, что эту регрессию мы можем отметить и на уровне отдельных клеток: имеется стрессовая де-дифференциация клеток, когда они становятся тотипотентными – они спускаются в дифференциации в такую точку, из которой потом могут образоваться разные типы клеток. Частично эта точка зрения используется в нашей концепции относительно канцерогенеза, которую мы недавно опубликовали в журнале «Медицинские гипотезы» (Medical Hypotheses, 2020). Мы считаем, что канцерогенез имеет отношение к регрессивному способу поиска новых путей организации клеточной активности в пределах целого организма. Как известно, исходя из представлений Давыдовского, патологические процессы – это не поломка нормального механизма, а выработанные в эволюции способы адаптации к среде, достижения положительных результатов в новых условиях, которые могут быть полезны для вида. Но не обязательно для каждого индивида они, эти попытки, заканчиваются позитивно. Мы знаем, что злокачественные клетки, опухоли образуются у всех в «нормальном развитии», потом апоптоз их убирает, но этот предохраняющий механизм срабатывает не всегда.
Эволюционное значение регрессии
Как могло в эволюции закрепиться такое контринтуитивное снижение дифференцированности актуализированного опыта в сложных ситуациях? Потому и закрепилось, что это, оказывается, адаптивно. Адаптивно для того, чтобы принять быстрое решение путем перехода к «древним», более простым и быстрым формам решения. Кроме того, как мы считаем, обратимая де-дифференциация может повысить эффективность действительно принципиально нового пути дальнейшего индивидуального развития.
Мы можем провести некоторые аналогии между структурами индивида, индивидуального опыта и структурами культуры. И те, и другие являются системными структурами; и там, и там действуют принципы селекции; и там, и там имеется переход к более дифференцированным системам (рис. 14).
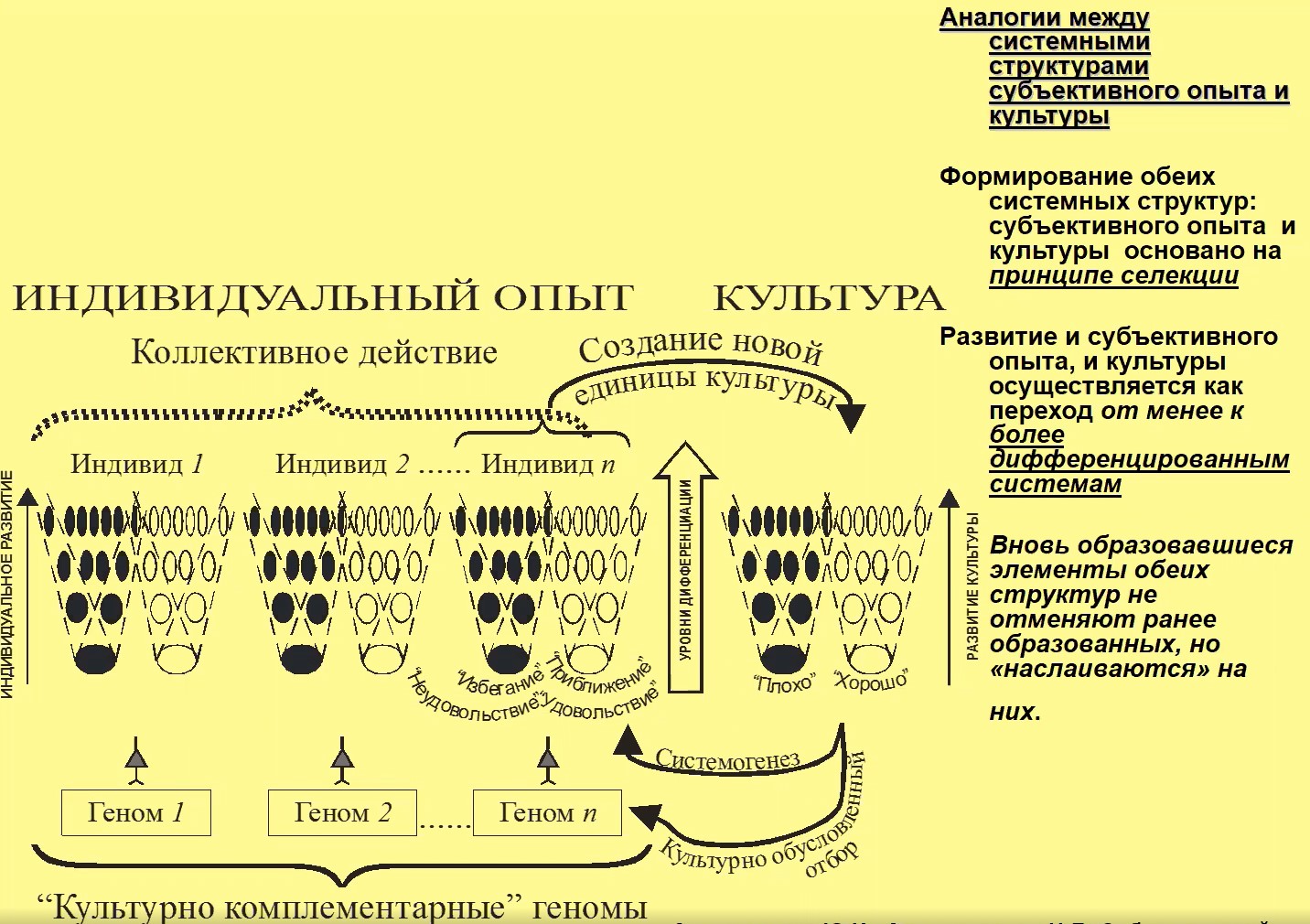
Рис.14
И соответственно с этим, и там, и там, действует регрессия – только про одну –«индивидную» регрессию мы уже рассказывали, а регрессия в культуре называется «архаизацией» (рис. 15). Авторы, изучающие архаизацию, говорят о том, что и она может способствовать дальнейшему развитию культуры.
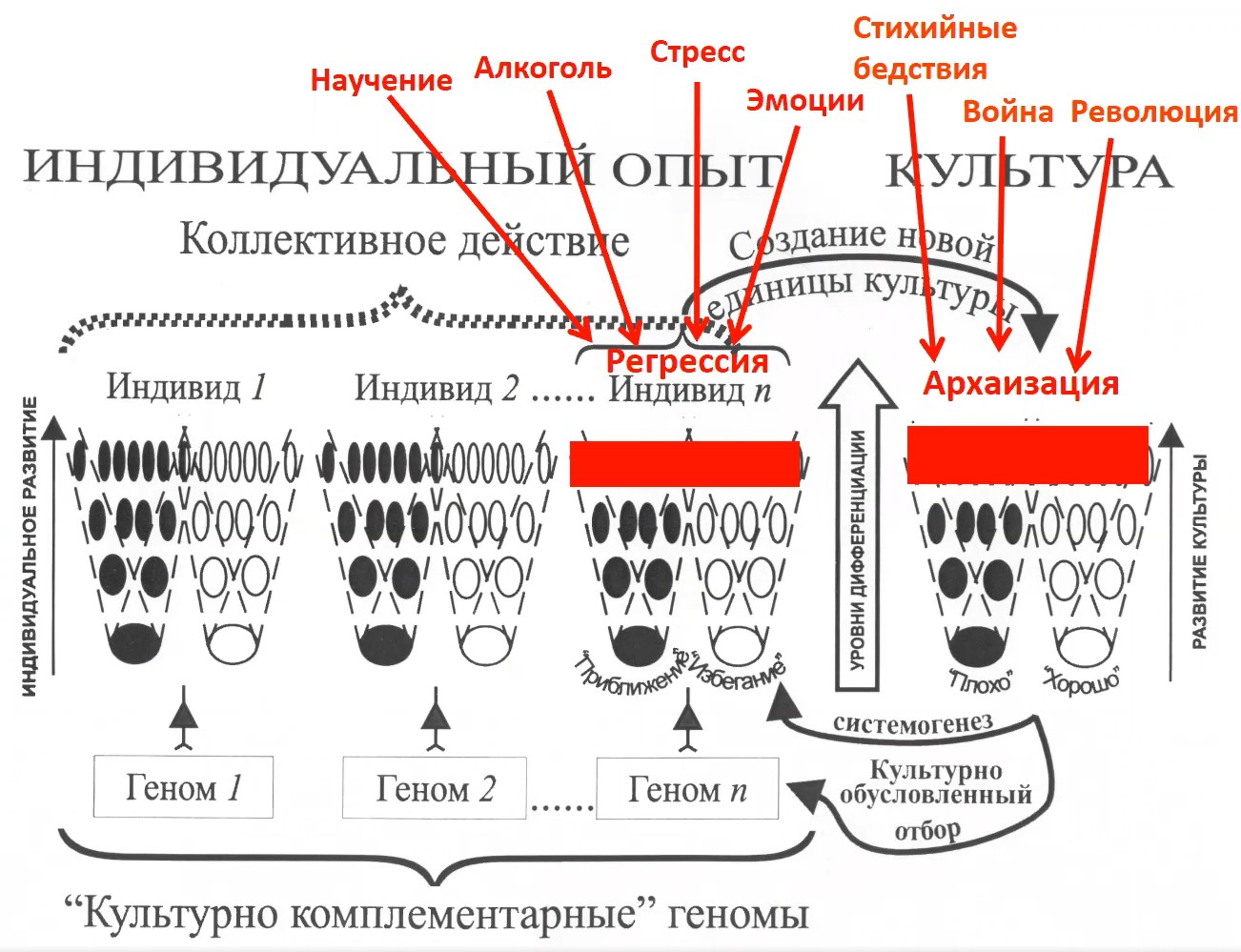
Рис.15
Когда мы говорим о различиях, то это различие касается не только нейронов или колоний, но и людей, оно выражается в различии их менталитета. Различие менталитета может касаться как отдельных культур, так и отдельных индивидов. Проявление различий менталитета может проявляться во время принятия решений по поводу моральной дилеммы, например, куда направить вагонетку: сбить одного, спасая пятерых, или допустить наезд на пятерых.
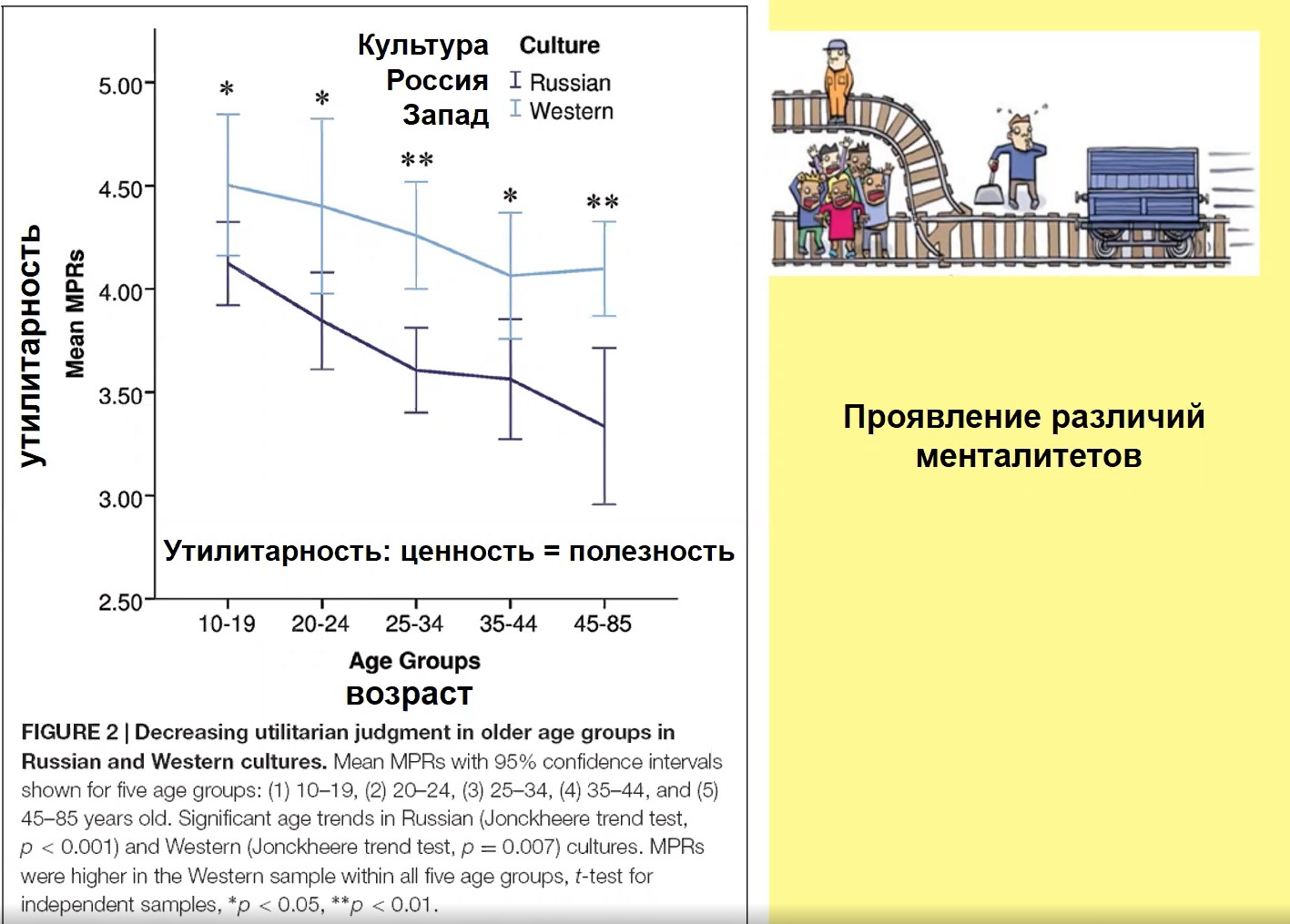
Рис.16
На рис. 16, представляющем результаты наших исследований, видно, что с возрастом люди все менее и менее готовы принять утилитарное решение, т.е. пожертвовать одним, чтобы спасти пятерых. Это уменьшение происходит как в англоязычных культурах, так и в России. Но заметьте, что в каждой точке, несмотря на общность этой тенденции для русской и «западной» культур, это решение всегда, в любом возрасте чаще принимается в англоязычной культуре. Иначе говоря, утилитарность в достоверно большей степени свойственна англоязычным культурам по сравнению с русской.
Посмотрите на карту индивидуализма и коллективизма (рис. 17, левый фрагмент):
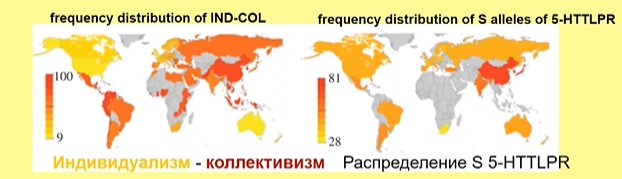
Рис.17
Видно, что эти карты соответствуют картам распределения гена серотонинового транспорта (рис. 17, правый фрагмент). Существуют разные аллели этого гена, и в коллективистских культурах чаще встречается короткий аллель, который ассоциирован с повышенной частотой отрицательных эмоций, вниманием к негативной информации, повышенным риском депрессии. Коллективизм может служить «защитой» от психопатологии («анти-психопатологическая функция»), которой в большей степени подвержены индивиды с коротким аллелем.
Ген-культурная ко-эволюция, связанная, в частности, и с описанной выше связью генома и ментальности, рассматривается в рамках расширенной теории эволюции, в которой не просто как в старой схеме среда оказывает влияние на генетический пул и отбирает соответствующие гены, которые попадают под действие следующей среды. Здесь (правый фрагмент рис. 18), при новом понимании эволюционного процесса, гены «строят» среду, и эта среда потом отбирает новые гены, которые строят новую среду, т.е. здесь двусторонние связи.
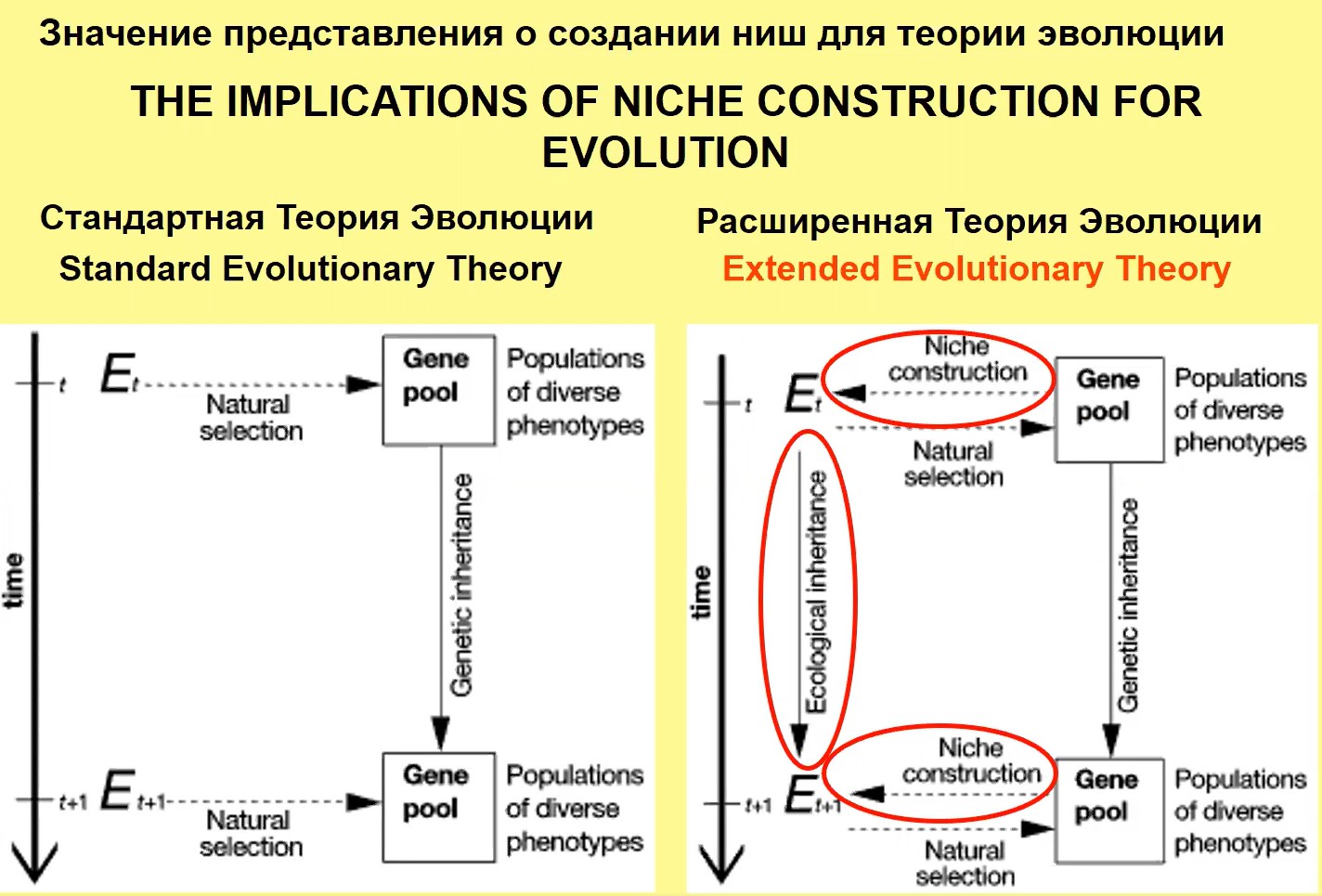
Рис.18
Формулировка этих представлений является одним из наиболее важных достижений в теории эволюции в XX веке. Имеется масса данных, которые показывают, например, как развитие молочного животноводства или определенных аграрных способов хозяйствования приводит к изменению генетической структуры популяции. Таким образом, те или иные способы хозяйствования связаны с теми или иными способами генетического обеспечения организма.
Такие передачи связаны с эпигенетическим наследованием. В настоящее время уже убедительно показано, что если мы учим животных определенному поведению, то это обучение связывается с изменением в ДНК с помощью метилирования определенных участков ДНК. Новое свойство ДНК передается не только первому поколению от обученных самцов, но даже второму поколению, которое произошло от наивных, необученных этому поведению самцов первого поколения, но все равно их потомки обладает предрасположенностью к выученному предками поведению.

Рис. 19
Здесь (на рис. 19) даны собранные мной симптомы, составляющие «западный» и «незападный» синдромы, которые представляют разные свойства соответствующих менталитетов. Важно, что здесь оказывается связанным индивидуализм с аналитизмом, а холизм с коллективизмом. Преобладание аналитизма свойственно западным европейцам и, особенно, американцам США, преобладание холизма – восточным азиатам, русским, китайцам.
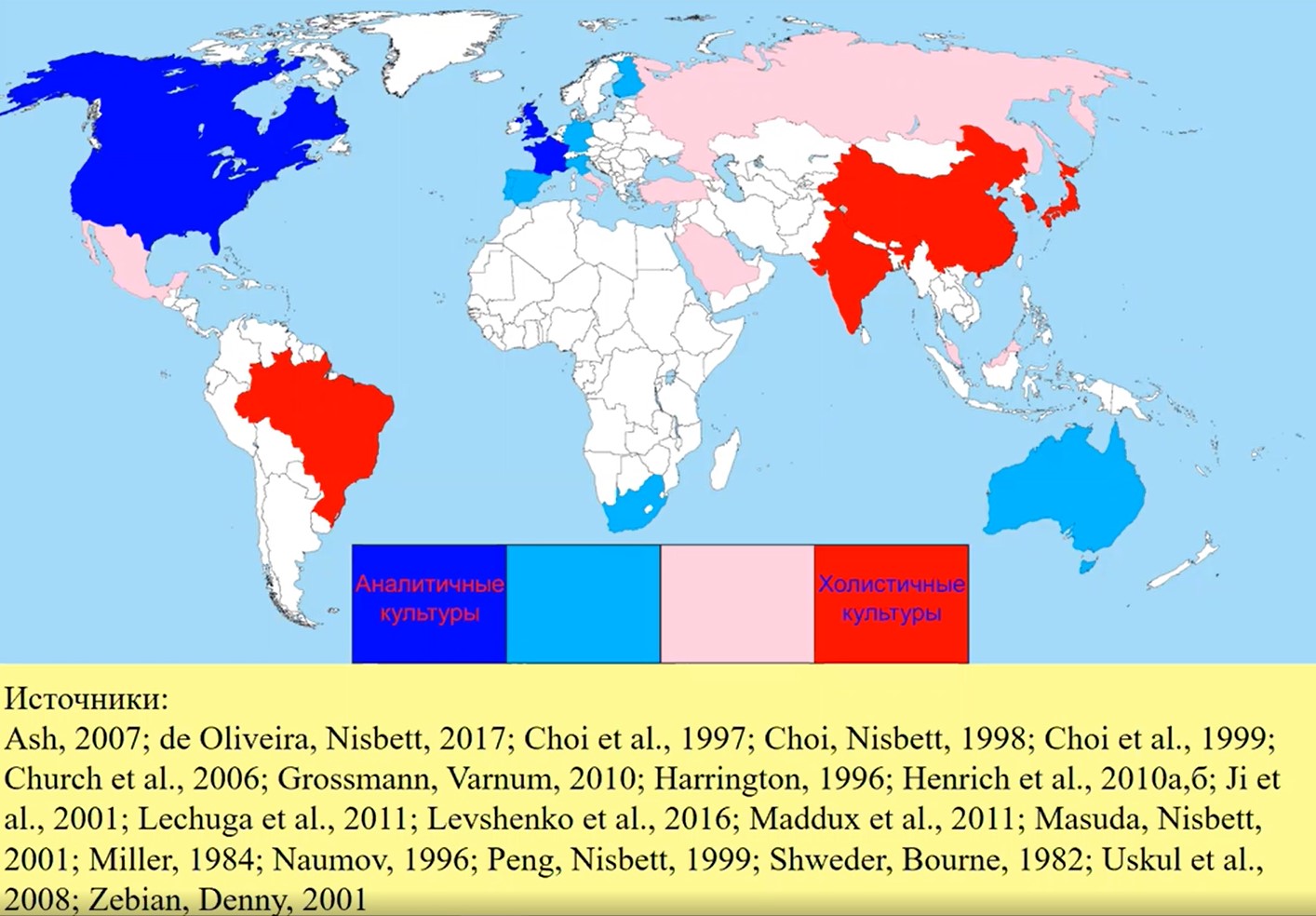
Рис.20
На рис. 20 представлены карты аналитизма и холизма, составленные В.В. Апановичем на основе данных целого ряда авторов. Эти карты интересно сравнить с представленными выше картами индивидуализм-коллективизма.
Индивидуализм и холизм – это не просто ментальные характеристики, они связаны с особенностями экономического, административного устройства общества и свойственны соответствующим, уже сопоставленным выше группам стран.
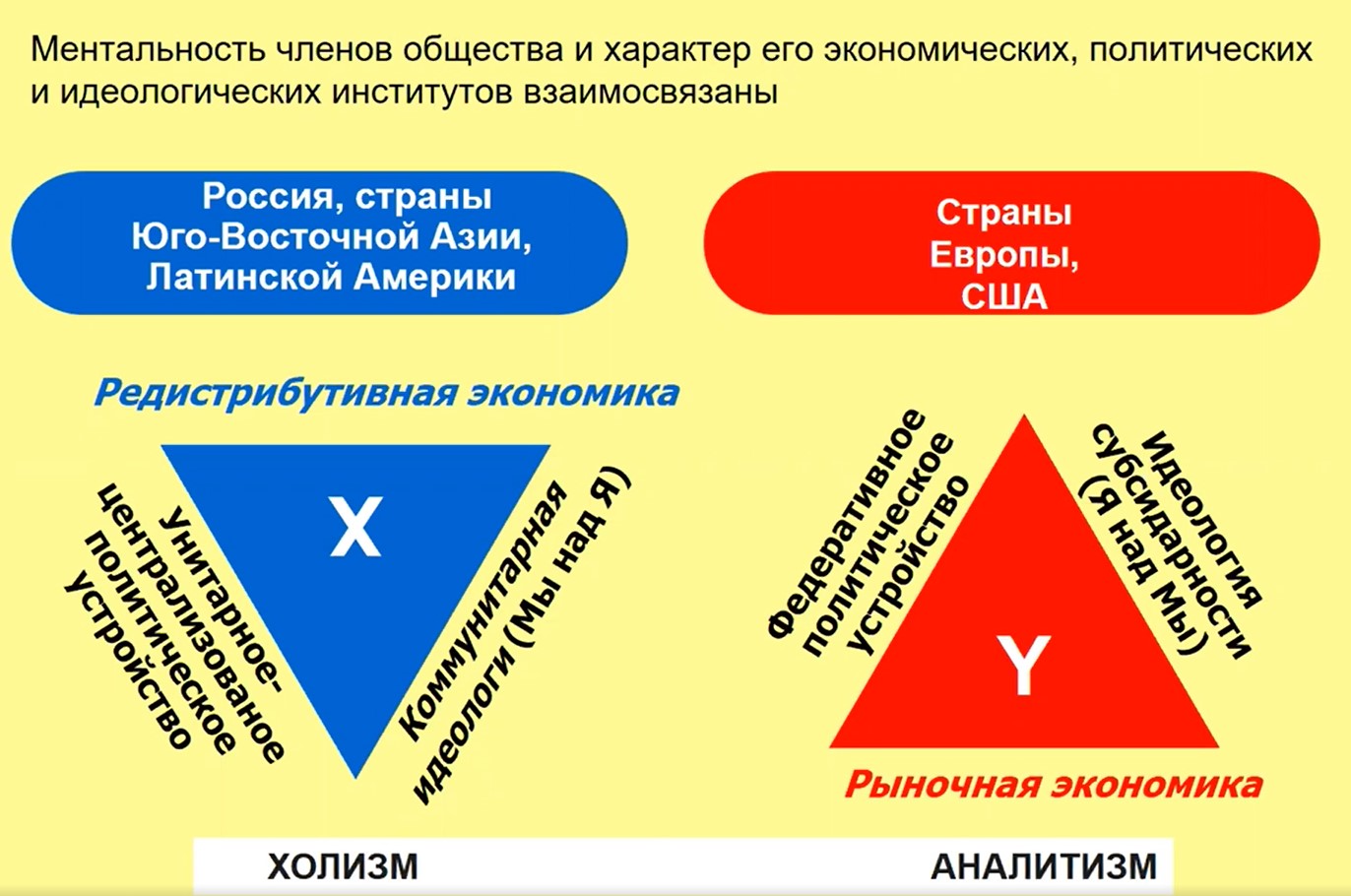
Рис.21
На рис. 21 представлены разные культуры, которые могут быть описаны экономикой, психологией и т.д. И эти культуры комплементарны. Нильс Бор писал, что «разные человеческие культуры дополнительны друг к другу, в отличие от физики здесь нет взаимного исключения черт, принадлежащих разным культурам». В каждой из культур присутствуют в качестве дополнительных, менее выраженных и свойства другой.
«Память культуры не только едина, но и внутренне разнообразна. Это означает… наличие частных «диалектов памяти», соответствующих внутренней организации коллективов, составляющих мир данной культуры», – писал Ю.М. Лотман. Эти «частные диалекты» могут быть обнаружены, например в Турции (см. рис. 22): здесь есть пастухи, рыбаки, земледельцы, и у них (а также в их семьях) разные типы менталитета, связанные с более и менее коллективным характером их труда. Мы можем также в Китае выделить «пшеничные» и «рисовые» области: «рисовые» области оказываются более холистичными (коллективистскими), а «пшеничные» более аналитичными (индивидуалистскими). Это связано с типом производства пшеницы и риса.
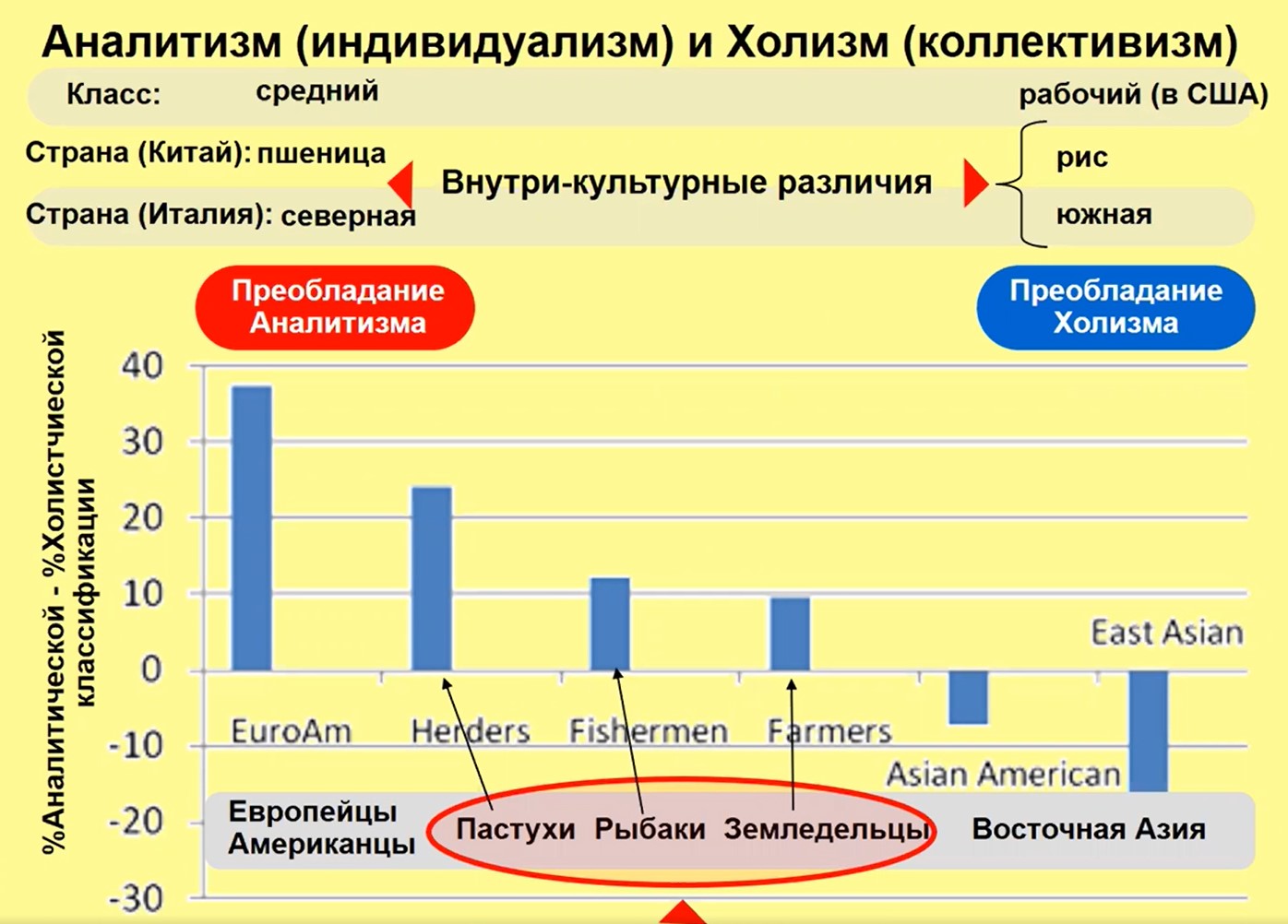
Рис.22
Наши исследования, проведенные совместно с финскими коллегами на финских субъектах (рис. 23), показывают, что у «холистов» и «аналитиков», ринадлежащих к одной культуре, мозг работает по-разному при совершении ими внешне одного и того же поведения.
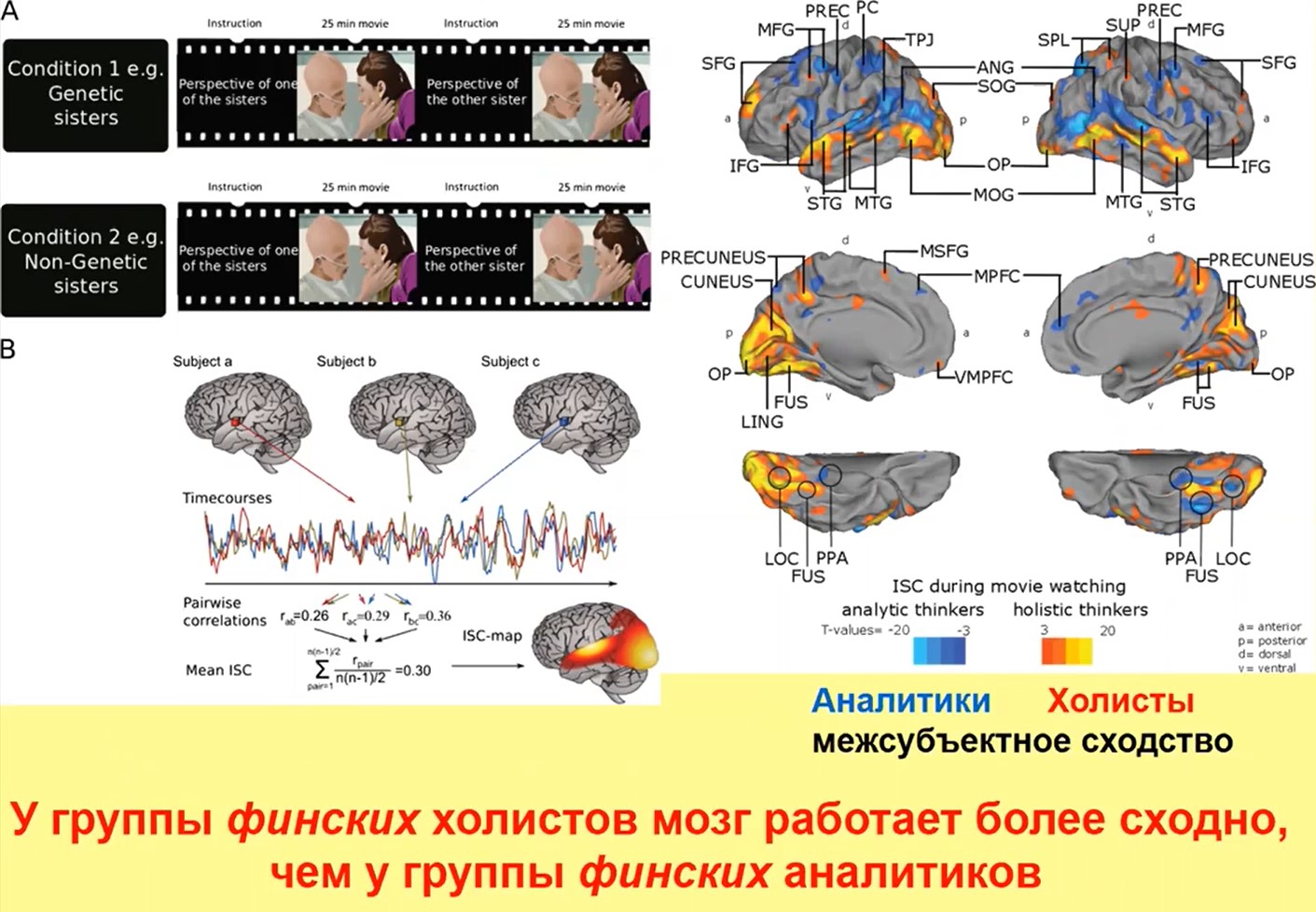
Рис.23
И на русской выборке мы обнаруживаем , что активность мозга при решении данной задачи по-разному связана у российских холистов и аналитиков с типом социальной ситуации, в которой решение происходит. Так, в ситуации конкуренции максимальная вариативность (при сравнении с ситуациями кооперации и индивидуального поведения) определенного компонента ЭЭГ-потенциала наблюдается у холистичных субъектов , но минимальная вариативность – у аналитичных субъектов (рис. 24). Отношение к типу социального взаимодействия у лиц с разным типом ментальности оказывается разным.
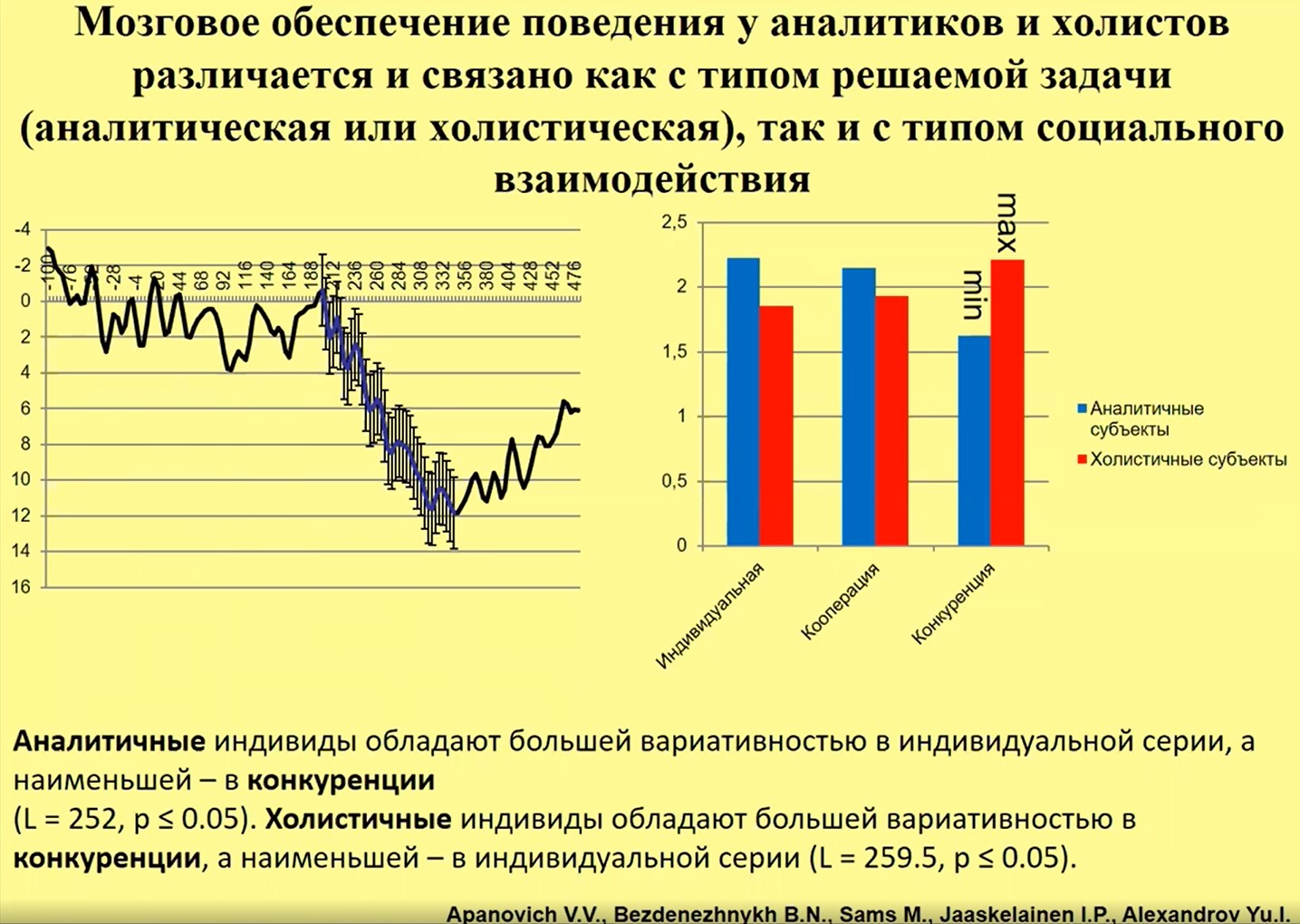
Рис.24
Наше математическое моделирование (А.К Крылов и соавторы) показывает, что в ситуации нестабильной среды кооперативность оказывается более выгодна для сохранения популяции, чем не кооперативность. Фенотипическое разнообразие тоже увеличивает выживаемость популяции (определяемую по ее численности), но кооперативность работает сильнее, чем разнообразие.
Связь среды и типа ментальности отмечена Нисбеттом: показано, что в соответствующей экологии развивается соответствующая форма хозяйствования, социальные практики, отсюда, как это уже было нами раньше отмечено, – соответствующая ментальность (рис. 25).
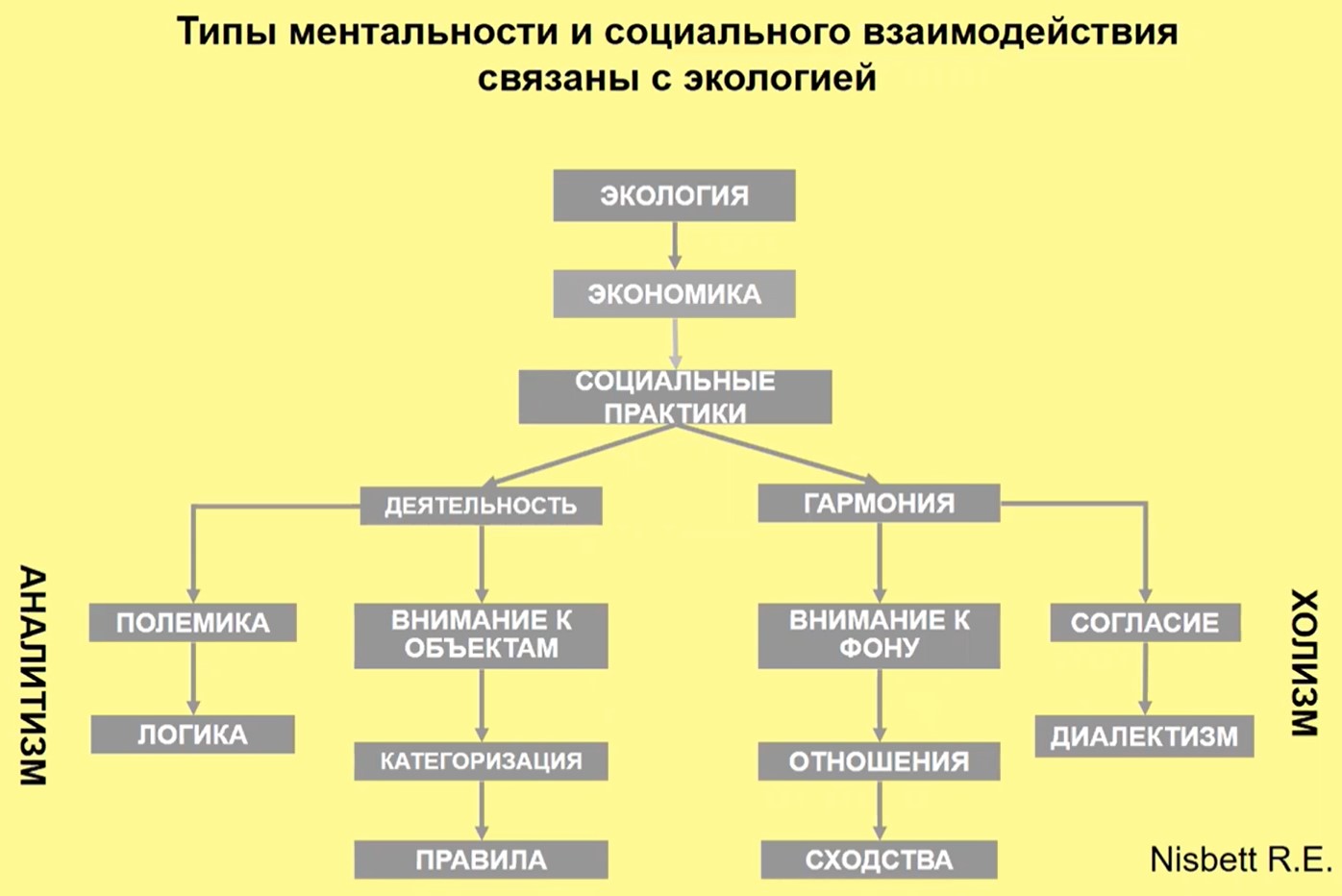
Рис.25
Могут быть выделены холистическая и аналитическая стадии познания (1 Х и 2 А на рис. 26). Гейзенберг говорил, что на одной стадии (1 Х) мы скорее чувствуем, чем анализируем, а на второй (2 А) начинаем анализировать и аккуратно собирать данные.
.jpg)
Рис.26
Если мы посмотрим на этот акт познания (рис. 27), то мы увидим, что холистическое мышление нам полезнее, эффективнее на первой стадии, аналитическое – на второй. Таким образом, холистическое и аналитическое мышление оказываются комплиментарными в общем процессе познания. Подчеркну, что это касается процесса познания как во внутрикультурном, так и в межкультурном смысле.
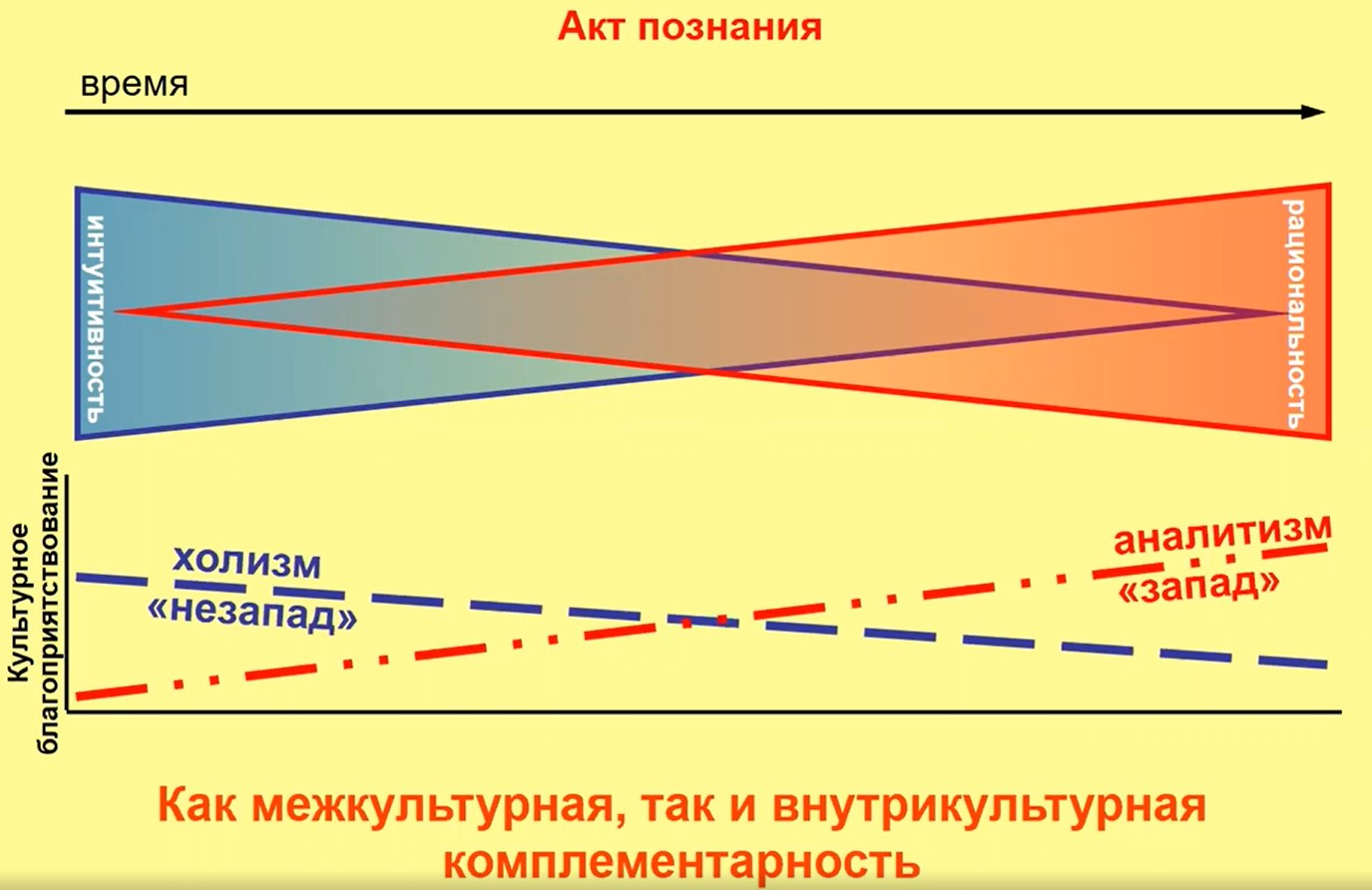
Рис.27
Действительно, нами получены данные, которые позволяют сказать, что аналитические и холистические задачи быстрее решаются в том случае, когда решающая эти задачи группа составлена из предположительно комплементарных пар (т.е. из аналитического и холистического субъектов), нежели чем из гомогенных пар. Я думаю, это пока очень гипотетически, что в этих парах регрессия при столкновении с новой ситуацией может происходить очень интересным образом: она будет проходить симметрично (выраженная регрессия у аналитиков и холистов, разводящая их на полюса аналитико-холистического континуума) либо асимметрично (больше выражена у тех, чья ментальность соответствует типу решаемой задачи)».






















































Комментариев пока нет – Вы можете оставить первый
, чтобы комментировать